減数分裂の制御における雌雄性差のメカニズム
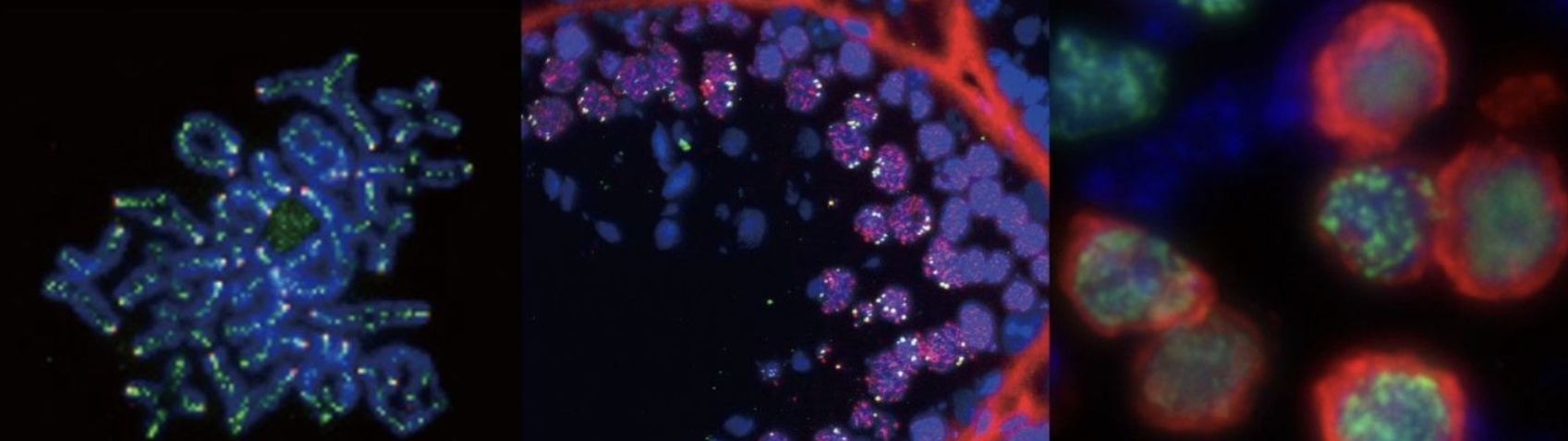
生殖発生のモードに応じて、減数分裂には雌雄で異なる調節機構が存在することが知られている。精巣においては「パキテンチェック」と呼ばれる監視機構により、減数分裂のエラーやトランスポゾンの異常活性化を検知し、異常な細胞を排除する仕組みが働いている。一方、卵巣では胎児期のごく短い時期に減数分裂が開始され、その後、減数第一分裂前期の途中で出生前にいったんDictyate arrestと呼ばれる停止状態に入る(図5)。この停止状態は、「diplotene stage の特殊な延長状態」とも見ることができ、ヒトでは、この休眠状態が数十年以上にわたって持続する。この停止状態の卵子はその後、徐々に限定された数だけ排卵にともない減数第一分裂を再開する。したがって卵子には減数第一分裂前期の進行を長期に渡って抑制するメカニズムが存在することが推定されるが、減数分裂における雌雄性差の本質的なことはあまりわかっていない。
我々はこれまで、遺伝子改変マウス、scRNA-seq、ChIP-seq、質量分析を組み合わせた研究を通じて、雄に特異的な減数分裂期の遺伝子発現制御様式を明らかにしてきた(Horisawa-Takada et al., Nat. Commun. 2021; Alavattam et al., Genes Dev. 2024; Yoshimura et al., Nat. Commun. 2024)。さらに、雌の卵母細胞と雄の精母細胞の減数第一分裂前期におけるscRNA-seqの比較解析から、雌に特異的に発現する候補遺伝子群を特定している。本研究では、これら雌雄特異的発現パターンを示す遺伝子の欠損マウスを用いた解析を通じて、生殖細胞の発生の文脈における減数分裂の雌雄差を調節するメカニズムの実態解明を目指す。
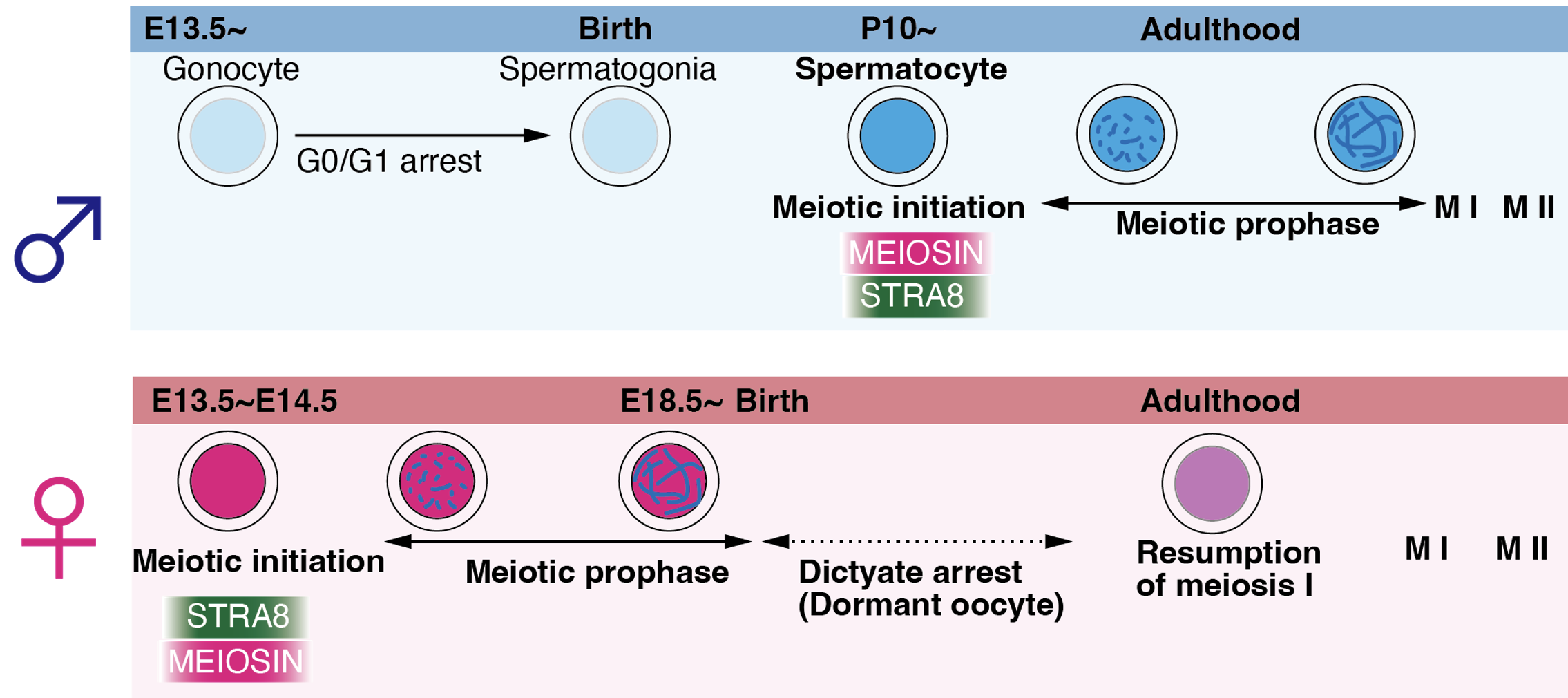
図5 減数分裂の制御における雌雄性差